应用岛屿生物地理学规划行星开发与保护
应用岛屿生物地理学规划行星开发与保护
Rubbish编辑部!!请切换编辑模式为分屏预览!!
作者名字1,*,†
1工作单位
*abc@example.com
†这些作者贡献相同。
乘煦
白鲸大学生态学院
ecoevo@beluga.edu.cn
摘要
生命所赖以生存的地球可能在未来变得不再宜居,寻找并开发具有潜在宜居环境的行星是应对这一危机的可能解决方案。20世纪60年代以来,岛屿生物地理学理论受到广泛的认可,并被应用在保护地理、入侵生物防治等方面。将岛屿生物地理平衡理论应用于描述行星上生物群落的动态变化是该经典理论在天体生物学领域的推广尝试。本文试图将浩瀚太空中的人类航道视作空间海洋、将行星视作太空岛屿,行星上的人类改造区视为亚岛屿,通过类比岛屿上的生物地理过程,描述一定环境条件下行星上生物群落的动态变化,并探究该过程在行星可承载物种数估计、行星生态系统的建立、维持与保护、星际生物入侵的管理等应用中的可能影响,为规划实施未来星际殖民计划提供生物地理学的视角。
关键词:岛屿生物地理平衡理论;天体生物学;行星开发与保护;入侵物种管理;系统保护设计
1 引言
许多研究表明,随着大陆漂移、气候变化、资源耗竭、生物多样性丧失等过程,我们现已知的全部生命所赖以生存的星球可能变得不再宜居。Xu等(2020)通过构建人类的气候生态位并将其与现今碳排放轨迹下预测的未来气候情景进行比较,认为50年后将有19% 的陆地区域亦即35亿人的居住地将超出人类适宜生态位,难以维系人类生存;Kang和Eltahir(2018)预测受气候变化和灌溉农业促发的极端热浪影响,2070-2100年华北平原可能不再宜居;O’Callaghan(2023)通过模拟2.5亿年后的地球系统,发现当各大陆重新聚合形成超级大陆后,92% 的地球表面将不再适于哺乳动物生存。面对这个充满悲观色彩和不确定性的未来,人类必须做好最坏的打算,寻求在危机迫近甚至爆发的背景下,延续地球生态系统的解决方案。

寻找并开发具有潜在宜居环境的行星是可能的出路之一。通过星际殖民计划使地球生态系统拓殖到其他行星,为延缓资源、环境矛盾的爆发赢得空间和时间。现有太空殖民相关的研究多停留在地外基地及生命维持系统的开发设计上(Edwards, 2021; Panesar & Ashkan, 2018; Ghidini, 2018),少有从管理生态学、生物地理学、入侵生物学等视角出发,探究行星开发和保护过程中生命扩散、定殖、保护、绝灭等问题的研究。
岛屿生物地理学是综合上述研究方向的可行理论框架。岛屿生物地理学理论由Robert MacArthur 和 Edward O. Wilson 于 20 世纪 60 年代首次系统提出(MacArthur & Wilson, 1963; 1967),并迅速发展成为众多交叉领域中的基础生物地理量化模型。MacArthur 和 Wilson 在专著 The Theory of Island Biogeography (1967) 中强调了研究岛屿生态系统的特殊价值和普遍意义,提出了岛屿生物地理平衡理论(Equilibrium Theory of Island Biogeography, ETIB),并对拓殖和扩散过程进行模型化。岛屿生物地理平衡理论是岛屿生物地理学理论的核心模型。模型认为物种灭绝速率与岛屿面积呈负相关,与现存物种数呈正相关;物种迁入率与种源距离呈负相关,与现存物种数呈负相关。迁入率大于灭绝率时,物种数有增长趋势;灭绝率大于迁入率时,物种数有降低趋势;而当迁入率等于灭绝率时,岛屿生物种数达到动态平衡,此时现存物种数即该岛屿在一定地理条件下可承载物种数的估计值。岛屿生物地理学的平衡理论,为解释宏观群落生态过程提供了定量模型,同时在生物多样性保护和保护地设计管理等领域具有重要应用价值(阎恩荣等, 2022)。
本文尝试通过类比岛屿上的生物地理过程,初步构建简单的行星生物群落动态分析框架,为规划实施未来星际殖民计划提供生物地理学的视角。
2 理论基础
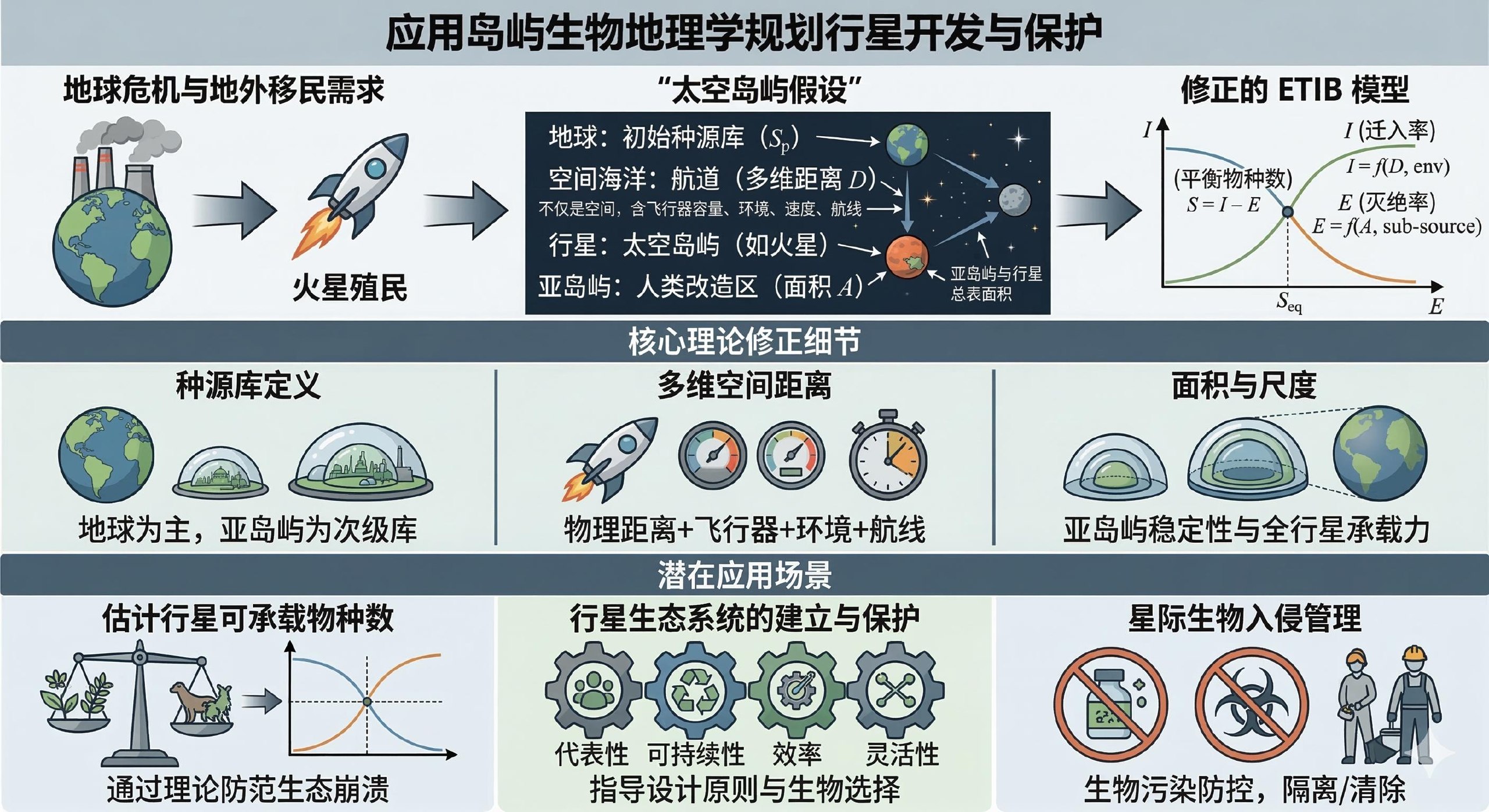
2.1 太空岛屿假设
岛屿生物地理学的观点和结论是否可以推广到星际生物关系呢?想要回答这个问题,首先需要考察宏观生物群落的栖息地——行星或岛屿——是否可以视为具有相似的性质。直观上看,行星具有由地理边界划定的有限面积,行星间具有难以逾越的地理障碍,行星上的生物有其种源星球、其星际扩散受距离衰减制约、一旦定殖到某颗空白星球则迅速占据可占据的生态位,似乎可看作漂浮在宇宙海洋中的岛屿。实际研究中也往往把具有类似性质的生境视作岛屿处理,如破碎化的栖息地(Simberloff & Abele, 1976; Harris, 1984)、空岛(Brown, 1971; Warshall, 1995; McCormack et al., 2009)。MacArthur 和 Wilson (1967) 也认为,岛屿生物地理学理论可以推广到孤立的栖息地,如“溪流、山洞、长廊森林、潮间带积水坑、苔原中的针叶林、针叶林中的苔原”。
但是,这种类比未免有些主观臆断了。实际上,将类岛屿系统直接视作岛屿并不恰当(Itescu, 2019)。Itescu 综述了各种各样类岛屿系统的特征,如空间破碎化、范围限制、时空隔离、被环境基质环绕等;然后考察了岛屿生物地理学理论被运用于这些系统的具体方面,如物种-面积关系、物种-隔离度关系、种群组成、辐射演化、遗传特有性和多样性等;最后构建了研究类岛屿系统的概念网络,并认为类岛屿系统具有宽泛的内涵,相比于真正的岛屿具有更低的隔离度,类岛屿系统中的生物模式只是部分地类似于真正的岛屿。Laurance (2008) 也从多个方面论证了在破碎化栖息地研究中应用岛屿生物地理理论的不足之处。
此外,即便行星可视作岛屿系统,若要将岛屿生物地理平衡理论应用于行星,必须对其进行一定修正。首先,一个星球的面积和环境异质性远大于一个岛屿,同时就目前可预见的科技进展来看,星际移民过程中生物的拓殖可能仅限于经人类改造过的星球表面,所以,不能直接将整个星球视作一个岛屿。我们可以将星球上经人类改造过的、适宜地球生物生存的区域视作亚岛屿,将一个星球上全部亚岛屿组成的集合视为一个大的岛屿。亚岛屿的总“面积”,既由星球本身的表面积和环境所决定,又受到人类对该星球的开发程度影响。其次,与少数生物可以跨过海洋拓殖到其他岛屿上不同,地球生物除非借助人造航天器运输,否则没有跨越宇宙空间的可能。所以,我们不能直接将星球间的物理距离视作“岛屿”间的距离,而要将与人类航道相关的因素纳入“距离”的考量。从星球内部的微观尺度看,我们还需要另外考虑同一星球上不同亚岛屿间距离的特点。
由此,我们提出太空岛屿假设:行星可视作太空岛屿,行星上的人类改造区可视为亚岛屿,行星间的人类航道可视作空间海洋,其中生物群落的动态变化符合岛屿生物地理平衡理论。由于技术限制,人类尚未在宇宙中发现生物星际拓殖的现象,也未在其他星球上进行生物地理学实验,所以,上述假设是完全空想而缺乏事实证据的。但是本文仍将试图从这一假设出发,推测星际生物地理动态的可能规律及潜在应用,为未来的太空探索和太空生物地理学实验提供可探究并验证或证伪的方向。这或许对生物地理学的发展具有些许启发意义。
2.2 太空岛屿上的岛屿生物地理平衡理论
太空岛屿上的岛屿生物地理平衡理论,需要从经典岛屿生物地理平衡理论推广得到。岛屿生物地理平衡理论反映了种源库距离和岛屿面积这两个地理因素,是如何通过迁入和灭绝这两个生态过程的平衡,影响岛屿物种数这一生物地理格局的。
公式1中, 为物种数, 为时间, 为迁入率, 为灭绝率。当 与 相等时, 趋于定值,即为岛屿平衡物种数。
迁入率与物种数 呈负相关,与种源库物种数 呈正相关,与种源库距离 呈负相关。灭绝率与物种数 呈正相关,与岛屿面积 呈负相关。因此,最终的平衡物种数取决于 、、、 四个变量。让我们在太空岛屿的假设下分别考虑这四个因素受到的影响和需要做出的修正。
2.2.1 物种数和种源库物种数
鉴于人类尚未在除地球外的其他星球上发现明确的生命迹象,我们姑且将地球视作唯一的初始种源库,地球生物圈总体物种丰富度对 有很大的贡献。此外,在拓殖某颗星球前已被拓殖的其他星球也可作为目标星球的种源库。
在目前已探明的、地球以外的星球上,只有被人类改造过的、接近地球环境的区域才能被多数生物拓殖,可视为星球内部的亚种源库。因为某一人为改造区域的环境由人类改造该区域时设定,该区域的环境异质性较低,环境相对单一,符合经典岛屿生物地理学理论对单一岛屿模型的要求。亚种源库是新建立的亚岛屿的潜在拓殖物种来源。一颗星球上亚岛屿物种数之和等于该星球的总物种数。
对于特定亚岛屿来说,其他星球作为一个整体向该亚岛屿输出新物种,而同一星球上的其他亚岛屿虽也能作为该岛屿的种源库,但并不影响该亚岛屿的灭绝率。因此,在针对某一亚岛屿进行计算时,物种数应选取该亚岛屿的物种数,而种源库物种数应选取种源星球的总体物种数或同星球其他亚岛屿的物种数。实际操作过程中,由于亚岛屿和亚岛屿之间的环境差异较大,可能无法用同一个物种平衡模型表示,计算平衡物种数时需要加入环境因子进行修正。
2.2.2 种源库距离
依前文所述,生物在星球间的扩散几乎只能以人类航道为通道,而且物种在特定通道的周转速率并不像经典岛屿生物地理平衡理论中的那样只取决于空间距离。星际飞行器的容量、环境、速度、航线长度等都是通道周转速率的影响因子。因此, 实际上是一种多维空间距离,以上述各影响因子为多维空间的维度。
2.2.3 岛屿面积
在考察太空岛屿面积时,必须单独考虑特定亚岛屿的面积,因为其他亚岛屿并不直接影响该亚岛屿的灭绝过程。但是,行星总表面积决定了该星球上能够建立多少、多大面积的人类改造区,因此,当整体考察一颗行星所能容纳的物种数时,行星总面积也是影响因素之一。
综上所述,太空岛屿生物地理平衡主要做出三点修正:(1) 针对某一亚岛屿进行计算时,物种数应选取该亚岛屿的物种数,种源库物种数应选取种源星球的总物种数或同星球其他亚岛屿的物种数,计算平衡物种数时需加入环境因子修正项;(2) 距离为星际飞行器的容量、环境、速度、航线长度等维度组成的多维空间中的距离;(3) 计算某一亚岛屿平衡物种数时,单独考虑该亚岛屿的面积,计算星球总平衡物种数时,行星表面积有一定影响。
3 行星开发与保护中太空岛屿生物地理平衡理论的可能应用
3.1 行星可承载物种数估计
由太空岛屿生物地理平衡理论可知,某一行星所能承载的物种数是有上限的,取决于行星表面积、亚岛屿数及面积加和、多维空间距离等客观因素。在设计星际移民总体规划时,必须维持物种数和生物个体数在阈值以下。一旦超过阈值,行星上的生态系统可能发生崩溃,总物种数向平衡物种数方向移动。
3.2 行星生态系统的建立、维持与保护
行星生态系统规划需要借鉴系统保护设计的四项原则,即代表性、可持续性、效率、灵活性。拓殖生物的选取应具有物种代表性,使天体生态系统具有复杂结构,增强稳定性;具有谱系代表性,使天体生态系统具有一定的抗风险性;具有功能代表性,使天体生态系统足以自发维持其生态功能的正常运作。规划需要实现可持续,各亚岛屿生态系统应能自我维持;亚岛屿间也应具有一定连通性,使群落有充足的生存空间。规划还应实现建设成本和成效之间的权衡,并具有充分的灵活性,才能使复杂的星际生态系统群体长期稳定可持续。这些原则的具体落实,都需要太空岛屿生物地理平衡理论预测结果的支持。
3.3 星际生物入侵的管理
当人类具有将生物送至其他行星的能力后,地球生物特别是地球微生物可能污染地外天体,其他天体的物质甚至地外生命也有污染地球的可能,这就催生了行星保护政策的诞生,该政策旨在采取有效措施避免地球和其他星球之间的生物和有机质污染,从而保护天体生物区并保证地外生命研究的可靠性(林巍等,2020)。
与普通生物入侵防治类似,星际生物入侵管理也需要从引入、建群、整合三个入侵步骤着手。由于星际飞行器是主要的引入通道,飞行器的生物安全就是引入段预防污染的重点。由于各亚岛屿之间是相对独立的,假若某一亚岛屿发生了污染,应当立即进行隔离,而后根据防治的可能性和成本的权衡做出继续隔离至清除污染或者解除隔离的决定,平衡物种数随各因素的变化是这一决定的重要参考信息。
4 前景展望
本文基于太空岛屿假设、经典岛屿生物地理平衡理论、星际拓殖的一些特点推出太空岛屿生物地理平衡理论的可能理论形式和潜在应用,为规划实施未来星际殖民计划提供生物地理学的视角。
必须再次强调的是,本文的全部推论都基于太空岛屿假设,而这一假设并没有实验依据。未来的天体生物学研究应尝试开展相关的野外实验,为探索星际生物地理学规律奠定实证基础。
参考文献
[1] 林巍, 李一良, 王高鸿, 等. 天体生物学研究进展和发展趋势[J]. 科学通报, 2020, 65(5): 380-391.
[2] 阎恩荣, 斯幸峰, 张健, 等. E. O. 威尔逊与岛屿生物地理学理论[J]. 生物多样性, 2022, 30(1): 22024.
[3] BROWN J H. Mammals on mountaintops: nonequilibrium insular biogeography[J]. The American Naturalist, 1971, 105(945): 467-478.
[4] EDWARDS M R. Android Noahs and embryo Arks: ectogenesis in global catastrophe survival and space colonization[J]. International Journal of Astrobiology, 2021, 20(2): 150-158.
[5] GHIDINI T. Regenerative medicine and 3D bioprinting for human space exploration and planet colonisation[J]. Journal of Thoracic Disease, 2018, 10(S20): S2363-S2375.
[6] HARRIS L D. The fragmented forest: island biogeography theory and the preservation of biotic diversity[M]. Chicago: University of Chicago Press, 1984.
[7] ITESCU Y. Are island-like systems biologically similar to islands? A review of the evidence[J]. Ecography, 2019, 42(7): 1298-1314.
[8] KANG S, ELTAHIR E A B. North China Plain threatened by deadly heatwaves due to climate change and irrigation[J]. Nature Communications, 2018, 9(1): 2894.
[9] LAURANCE W F. Theory meets reality: how habitat fragmentation research has transcended island biogeographic theory[J]. Biological Conservation, 2008, 141(7): 1731-1744.
[10] MACARTHUR R H, WILSON E O. An equilibrium theory of insular zoogeography[J]. Evolution, 1963, 17(4): 373-387.
[11] MACARTHUR R H, WILSON E O. The theory of island biogeography[M]. Princeton: Princeton University Press, 1967.
[12] MCCORMACK J E, HUANG H, KNOWLES L L. Sky islands[M]//GILLESPIE R G, CLAGUE D A. Encyclopedia of islands. Berkeley: University of California Press, 2009: 839-843.
[13] O’CALLAGHAN J. This is what Earth will look like in 250 million years[J]. Nature, 2023, 622(7981): 20.
[14] PANESAR S S, ASHKAN K. Surgery in space[J]. British Journal of Surgery, 2018, 105(10): 1234-1243.
[15] SIMBERLOFF D S, ABELE L G. Island biogeography theory and conservation practice[J]. Science, 1976, 191(4224): 285-286.
[16] WARSHALL P. The Madrean sky island archipelago: a planetary overview[R]//DEBANO L F, FFOLIOTT P F, ORTEGA-RUBIO A, et al. Biodiversity and management of the Madrean Archipelago: the sky islands of southwestern United States and northwestern Mexico. Fort Collins: U.S. Department of Agriculture, Forest Service, 1995: 6-18.
[17] XU C, KOHLER T A, LENTON T M, et al. Future of the human climate niche[J]. Proceedings of the National Academy of Sciences, 2020, 117(21): 11350-11355.
本博客所有文章除特别声明外,均采用 CC BY-NC-SA 4.0 许可协议。转载请注明来自 Rubbish!